时间:2020-09-08 15:00
1, AAV血清的选择
自然界中病毒感染细胞的过程大体是相似的,包括吸附、侵入、脱壳(有包膜的需脱包膜)、基因组复制和蛋白合成、组装和释放。通常情况下,感染的效率与病毒表面蛋白与细胞膜受体蛋白、多糖等分子之间的特异性识别相关。
AAV病毒家族成员被划分为多个血清型,常见的如表1,它们的衣壳蛋白表达谱并不完全相同,所以在感染细胞时会产生一定的偏好性。比如AAV1, AAV5, AAV8 和AAV9不仅能感染神经元,还能感染神经元周围的胶质细胞;而AAV2对神经元的感染效率高,对胶质细胞的感染性低。利用这个特点,当研究脑区特定神经元定位和投射时,选择像AAV2这类神经元特异性较强的血清型可降低非特异性感染的背景,提高信噪比,至少可以缩小血清型选择的范围。
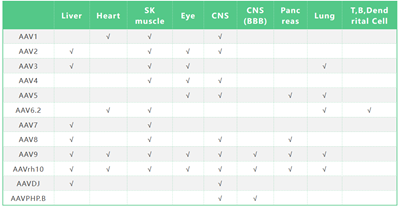
表1. 常见血清型AAV的组织感染嗜性
以往的研究发现,不同血清型AAV感染细胞的类型、效率、以及病毒入侵细胞后表现出的动力学特征存在一定的差异,研究者通过使用巨细胞病毒CMV强启动子表达荧光素酶标记系统的AAV对动物进行尾静脉注射,在不同时间点采集荧光信号,得到AAV在体内感染的动态分布图,可以评估每一个血清型在个体不同组织中表达的偏好性[1]。
2,给药/注射方式的选择
以研究神经系统为例,AAV给药方式通常有原位注射,静脉注射,和脑脊液注射三种。原位注射是最直接最有效的传导方式,所需剂量较少,靶向性强,但会存在局部传导的梯度逐渐降低的问题,比如脑区原位注射之后很难扩散到离注射点更远的脑深处。另外,如果操作不当容易引起动物的创伤性综合反应。具体的注射方式取决于具体的实验需求和病毒血清型特点。
血脑屏障是指脑毛细血管壁与神经胶质细胞形成的血浆与脑细胞之间的屏障和由脉络丛形成的血浆和脑脊液之间的屏障,这些屏障能够阻止某些物质(多半是有害的)由血液进入脑组织。一般情况下,即便是很小的化学分子都无法跨越过屏障,但存在部分经改造的engineered AAV,如AAV-PHP.B,能通过无创静脉注射经血液循环跨过血脑屏障进入脑部中枢神经系统,这似乎为研究脑神经系统疾病相关的药物分子代谢动力学提供了一些思路。
经过定向进化选择的方式筛选出的rAAV2-retro病毒株,能感染神经元轴突末端,有高效率逆向传输的活性。当研究中枢神经系统内多级连接环路时,比如想知道大脑的定点区域功能受上游哪些脑区、核团、神经元的调控,存在怎样的连接,则通过原位脑区注射rAAV2-retro等逆向跨突触传输的病毒来研究该环路。另外,也可通过其他类型重组嗜神经病毒(如PRV,HSV等)原位脏器注射,绘制中枢神经系统⇋外周神经系统特定的传入和传出解剖学结构和连接模式[2]。
到目前为止,血清型和给药方式的选择并没有公认的最佳方案,往往需要回顾与实验最相关的文献,结合实验室已有的经验,做必要的预实验对表达效果进行测试。表2例举三种不同注射方式下的血清型AAV的选择参考(神经系统)。

表2. AAV的给药/注射方式(左),血清型表达特点(右)
3,细胞毒性
尽管AAV相对其他病毒而言拥有更低的免疫原性,但并不意味着对细胞活动不造成影响,尤其在高剂量注射的情况下。研究发现向静脉、外周和中枢神经系统注射AAV均会引起神经毒性。因此,在开始全新的实验之前,需要结合动物的年龄、生理状态等条件设置注射剂量的梯度对照。一些研究表明AAV感染肌肉和肝脏时刺激TLR9而激活先天性免疫反应,设计含有阻断TLR9转录的寡聚核苷酸序列整合到的AAV载体图谱中,可减少免疫反应的程度。
1,多顺反子表达元件
顺式调控元件可用于控制AAV在一些模式生物和转基因哺乳动物中对特定细胞类型进行转基因表达。这些元件包括启动子和增强子(图1C),以及内含子、MicroRNA识别序列和可用于RNA剪切和翻译的内部核糖体进入位点IRES(图1D),以满足载体基因表达的需要。
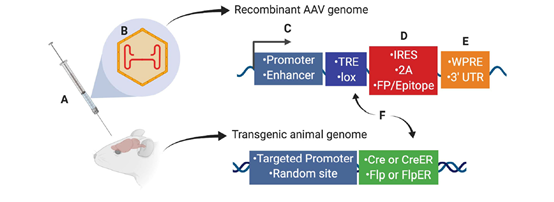
图1. AAV载体的顺式调控元件序列示例图,图源于文献3
然而,单个AAV基因组的包装能力有限,能携带外源基因片段长度一般不超过4.7 kb,因此,开发更短的启动子序列和设计多顺反子表达元件提高单个载体的空间利用率一直是热门的研究领域。可利用以下方法实现在一个载体上实现多个基因的共表达。
(1) 由于IRES元件具有不依赖帽子结构而独立募集核糖体的功能,进而可以启动下游基因的翻译(虽然翻译效率不及IRES上游),所以使用IRES 序列实现多顺反子的共表达;
(2)2A元件在翻译过程中形成的高级结构对核糖体肽基转移酶中心造成空间位阻,导致无法形成正常的肽链连接,但同时核糖体却能继续翻译下游蛋白,从而形成一种类似蛋白水解酶的作用将前后两个蛋白顺式“切开”,使用2A自切割肽序列能实现同一顺反子的分割表达。
实际运用中,比如需要令两个蛋白分开表达(例如一个表达荧光报道蛋白,另一个表达细胞膜受体蛋白),但受限于载体容量,只想在载体上构建一个开放读框时,则可在其编码区之间插入一段2A肽序列。
2,转录后的调控
转录后的调控对前体mRNA剪接、mRNA稳定性和出核、以及翻译都有非常重要的作用。AAV载体靠近3’端内含子上的WPRE元件,能增强载体的表达及转录后mRNA的稳定性,提高mRNA剪接效率。另外,在3‘端非翻译区可设计插入一些识别非靶向细胞中转录的miRNA互补核苷酸序列,降低载体在非靶向细胞中的表达率,提高了表达的细胞特异性。比如肌肉细胞转录miR-1,肝脏细胞转录miR-122,设计与miR-1和miR-122互补的核苷酸序列插入到AAV9的3‘端非翻译区,这样以来,当通过静脉注射AAV9感染动物时,便减少了AAV9在肌肉,心脏,肝脏组织细胞中表达几率,而保持着对靶向区域--大脑的特异性感染和表达。
3, 构建条件型诱导表达系统
哺乳动物的中枢神经系统包含多种不同类型的神经元,这些神经元各自拥有独立的基因表达谱,在生物体整个发育过程中受到各式各样的调节。通常情况下,基因的表达除了需要固有的顺式调控元件(启动子和增强子等基因序列),还需要反式作用因子参与调节,如RNA聚合酶及相关的转录因子,特异性激活子和抑制子等。反式作用因子的调控作用实际上是通过不同途径改变染色质和DNA空间构象,或搭建转录因子与基因序列之间互作的桥梁,确保基因的表达能正常进行。基于反式作用因子的调控原理,研究人员构建了基于四环素(TET)反应元件TRE和基于重组酶依赖的表达系统(如重组酶Cre能识别loxP位点),当我们想要实现AAV载体转染后的时空特异性表达时,这些策略似乎非常合适。
对于Tet系统,将Tet-on反向四环素反式激活子(rtTA)或Tet-off(tTA)与带有Tet响应元件(TRE)的启动子的AAV载体配对。比如在Tet-on调控系统中,当Dox(强力霉素)不存在时,rTetR不能与TRE结合,基因表达被抑制;而当Dox存在时,rtTA能与TRE结合,进而使得目的基因表达(图2C)。因此,Tet策略能实现对靶向基因表达时间点的控制,需要表达时给药诱导,不需要表达时则不给药,或去除诱导剂终止表达。
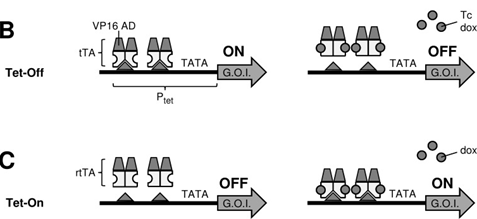
图2. Tet诱导系统,图源于文献5
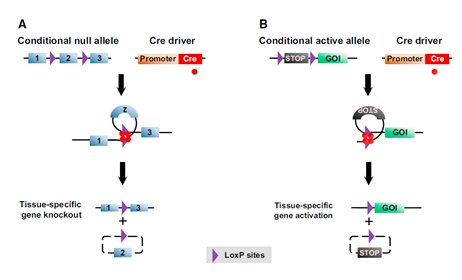
反过来,我们可以通过该种策略构建和筛选Cre和Flp定向且稳定表达的转基因小鼠系,设计Cre和Flp依赖的细胞特异性启动子表达开关的载体,控制开启效应子或报道基因的表达,实现对感兴趣的特定细胞的标记和操纵(图3B,Cre酶识别Lox-stop-Lox终止盒位点选择是否表达)。
另外还存在一类受tamoxifen(它莫西芬,一种雌激素受体分子药物)调节重组酶CreER和FlpER活性的更加精准的诱导体系,只有在tamoxifen存在的条件下,重组酶依赖的效应子或报道基因才能被调控。
总体来说,本篇文章仅对AAV载体在特定细胞类型中实现靶向基因的传递策略进行了初步的分析和探讨,然而对于具体实验中所遇到到的问题要更为复杂。实际上,AAV传递策略仍面临着很大的挑战,如何评估构建的载体和转基因动物的有效性,如何降低AAV载体制剂引起的细胞毒性和机体免疫反应,如何携带更多传感器、功能酶以及遗传编码工具实现更精准更高效率的表达等,都是研究者今后努力的方向。
【参考文献】
1,Carmela Zincarelli et al,. Analysis of AAV serotypes 1-9 mediated gene expression and tropism in mice after systemic injection Mol Ther. 2008 Jun;16(6):1073-80. doi: 10.1038/mt.2008.76.
2, Fan L, Xiang B, Xiong J, He Z, Xiang H, Use of viruses for interrogating viscera-specific projections in central nervous system, Journal of Neuroscience Methods (2020), doi: https://doi.org/10.1016/j.jneumeth.2020.108757.
3,Leila Haery et al,. Adeno-Associated Virus Technologies and Methods for Targeted Neuronal Manipulation. Frontiers in Neuroanatomy. 26 November 2019. doi: 10.3389/fnana.2019.00093
4,Magnuson M , Osipovich A . Pancreas-Specific Cre Driver Lines and Considerations for Their Prudent Use[J]. Cell Metabolism, 2013, 18(1):9-20. DOI: http://dx.doi.org/10.1016/j.cmet.2013.06.011
5,Atze T Das et al,. Tet-On Systems For Doxycycline-inducible Gene Expression. Curr Gene Ther. 2016; 16(3):156-67. doi: 10.2174/1566523216666160524144041.
香港宝典免费资料网提供各种类型的AAV
只要你想不到,没有我们做不到!
感兴趣的话快来联系我们吧~
本文未经授权禁止转载
如有需要,可联系小编获取原文